|
|
|
|
|
BOTANIK |
Die Lichtabsorbtion bei der Photosynthese |
|
 |
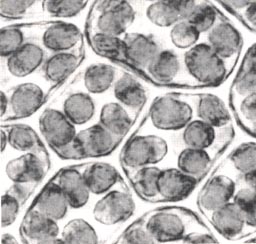
LichtabsorbtionBau der Chloroplasten
Die Chloroplasten höherer Pflanzen sind diskusförmig bis ellipsoid.
Ihr Durchmesser liegt zwischen 3 und 10 µm. Bei höchster lichtmikroskopischer
Auflösung erkennt man in ihnen ein Muster von dunkelgrünen Zonen
(Grana) auf hellerem Untergrund. Die BlattfarbstoffeIn den Tylakoidmembranen sind die Blattfarbstoffe (Blattpigmente) lokalisiert. Wenn man aus grünen Blättern Farbstoffe extrahiert, werden sie aus den Membranen herausgelöst. Trennt man die extrahierte Farbstofflösung chromatographisch, erkennt man grüne, gehe und orangefarbene Farbstoffe. Bei den gelben und orangefarbenen Farbstoffen handelt es sich um Carotinoide. Bei den grünen Farbstoffen kann man das dunkelgrüne Chlorophyll a und das gelbgrüne Chlorophyll b unterscheiden. Die unterschiedliche Wanderungsgeschwindigkeit der Farbstoffe hängt mit der Molekularstruktur und den verschiedenen chemischen Eigenschaften dieser Farbstoffe zusammen. Das Chlorophyll besteht aus vier miteinander verbundenen Pyrrolkernen, die durch ein System von abwechselnden Einfach- und Doppelbindungen miteinander verbunden sind. Diese konjugierten Doppelbindungen sind verantwortlich für die grüne Färbung. Am Pyrrolkern IV sitzt ein lipophiler Alkoholrest, das Phytol. Bei der chromatographischen Trennung ist dieser Rest zum Beispiel gut in Petroleumbenzin löslich. Daher wandert das Molekül entsprechend weit. Carotin besteht aus Isopreneinheiten mit zwei endständigen Ringsystemen, die besonders lipophil sind. Entsprechend wandert es noch weiter als das Chlorophyll. Blattfarbstoffe absorbieren Licht:
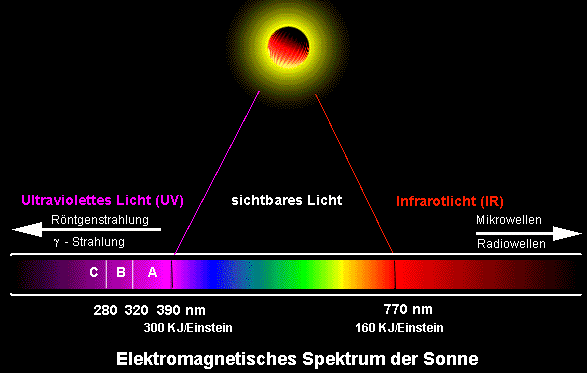
Abb.: Das Lichtspektrum der Sonne. Der sichtbare Bereich ist nur ein kleiner Teil des gesamten Spektralbereichs. Die Sonne ist die universelle Energiequelle, die für den Aufbau der Biomasse und den Betrieb der Lebensprozesse auf der Erde verantwortlich ist. Bei den in der Sonne ablaufenden Kernfusionsprozessen wird Materie in Energie umgewandelt. Diese Energie wird in Form elektromagnetischer Strahlung in den Weltraum abgegeben. Ein Teil dieses Spektrums wird von der grünen Pflanze absorbiert. Es handelt sich dabei um den Wellenbereich zwischen 380 und 780 nm. Will man genauere Aussagen über die Lichtabsorption verschiedener Pigmente machen, so bringt man sie in Lösung und bestimmt mit Hilfe eines Spektralphotometers für jede Wellenlänge die Absorption. Das Spektrum des Chlorophylls zeigt ehe starke Absorption im Bereich der roten und blauen Strahlung. Grüne und dunkelrote Strahlen werden dagegen kaum oder gar nicht absorbiert. 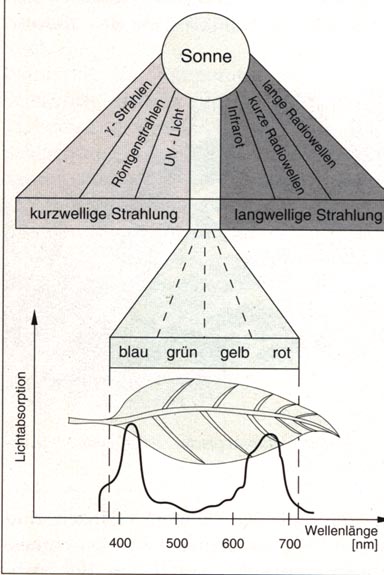
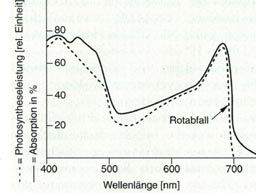
Abb.: Absorbtionsspektrum von Chlorophyll a. Der Absorptionsvorgang besteht in der Aufnahme dieser Strahlung. Die kleinsten absorbierbaren Einheiten der Strahlung werden Photonen oder Quanten genannt. Die Energie eines Lichtquants ist nicht konstant. Sie verhält sich umgekehrt proportional zur Wellenlänge der Strahlung. Je kürzer die Wellenlänge, desto höher seine Energie. Strahlung lässt sich also neben der Wellenlänge auch durch den Energiegehalt ihrer Quanten definieren. E = h * f E = h * c/l h = Plancksches Wirkungsquantum (6,6*10-34) Das langwelligere rote Licht ist weniger energiereich als das kurzwelligere blaue Licht. Bei der Aufnahme von Lichtquanten durch Atome oder Moleküle ändert sich deren energetischer Zustand. Um den Kern eines Atoms bewegen sich Elektronen in Orbitalen. Durch Lichtenergie angeregt werden sie schneller und können in ein kernferneres Orbital gehoben werden, d. h. auf ein höheres energetisches Niveau. Sie werden vom Grundzustand in den angeregten Zustand überführt. Nur ein Lichtquant, dessen Energiegehalt der Differenz zwischen diesen beiden Zustandsformen entspricht, kann Elektronen anregen. Im Chlorophyllmolekül haben die konjugierten Doppelbindungen Orbitale zur Folge, die dem gesamten Ringsystem gemeinsam sind. Im Molekül befinden sich daher besonders viele frei bewegliche Elektronen (Elektronen), die dem konjugierten System als Ganzem zugeordnet werden können. Durch Absorption eines Rotlichtquants wird das Chlorophyllmolekül in den sogenannten 1. Anregungszustand überführt. Durch ein blaues Lichtquant wird es in den 2. Anregungszustand gebracht, d. h. auf ein noch höheres Energieniveau. Bereits nach 10-12 s fallen die Elektronen vom zweiten in den ersten Anregungszustand zurück. Die Energiedifferenz wird als Wärme frei. Auch der erste Anregungszustand ist außerordentlich kurzlebig, nämlich 10-9 s. Diese Zeit reicht jedoch, um die Energie dieses Zustandes photochemisch wirksam werden zu lassen, d.h. um einen Elektronentransport einzuleiten. Ist dieser Transport nicht möglich, dann geht die absorbierte Energie als Wärme oder als Fluoreszenzlicht verloren oder wird auf ein Nachbarmolekül übertragen. Chlorophylle und Carotinoide sind zu photochemischen Reaktionseinheiten zusammengefasst. Man nennt sie Farbstoffkollektive oder Photosysteme. Sie sind eingelagert in die Thylakoidmembran der Chloroplasten und bestehen jeweils aus rund 300 Farbstoffmolekülen. Alle Farbstoffmoleküle können Licht absorbieren. In jedem System ist jedoch nur ein Molekül Photomechanisch wirksam. Es handelt sich immer um Chlorophyll a. Man bezeichnet es als Reaktionszentrum. Die übrigen Farbstoffe nennt man Antennenpigmente. Sie übertragen die von ihnen absorbierte Energie auf das Reaktionszentrum. Diese Energieübertragung geschieht durch Resonanz, d. h. die Energie wird durch Schwingungen nahezu verlustfrei übertragen. Dieser Resonanztransfer ist möglich, wem das benachbarte Molekül bei gleicher oder etwas größerer Wellenlänge absorbiert. Entsprechend besitzt sein Anregungszustand ein etwas niedrigeres Energieniveau. Die Energiedifferenz geht als Wärme verloren. Dieser Mechanismus ermöglicht nur den Transport einer Richtung, nämlich zum Reaktionszentrum hin. Modellhaft kann man sich ein Photosystem trichterförmig vorstellen. Das Licht trifft auf die obersten Farbstoffmoleküle und regt sie an. Sie leiten durch Resonanztransfer die Energie an die darunterliegenden Moleküle weiter bis hin zum Reaktionszentrum. Man bezeichnet diesen Mechanismus als "Lichtsammelfalle". Das Reaktionszentrum kann jeweils nur die Energie eines einzelnen Lichtquants aufnehmen. Während dieser Zeit ist es für die übrigen Quanten besetzt. Wenn man eine bestimmte Lichtmenge portioniert, d. h. nicht auf einmal sondern in kleinen aufeinanderfolgenden Lichtblitzen verabreicht, kann man mit der gleichen Menge eine bessere Quantenausbeute erreichen. Durch Portionierung des Lichtes lässt sich eine Steigerung der Photosyntheserate um das acht- bis neunfache erreichen. Die beiden Photosysteme
In den Bereichen oberhalb und unterhalb von 682 nm (Nanometer) ist die Wirksamkeit der in diesem Spektralbereich auftretenden Lichtquanten in Relation zu ihrer Absorption jeweils etwas verringert. Im Bereich unterhalb 682 nm liegt das daran, dass die von den Carotinen absorbierte Strahlung weniger effektiv verwertet wird. Die geringere Wirksamkeit oberhalb von 682 nm, der sogenannte Rotabfall, wird dadurch nicht erklärt. Bestrahlt man Chloroplasten mit Licht der Wellenlängen oberhalb von 682 nm und gleichzeitig mit kurzwelligerem Licht einer Wellenlänge unterhalb von 682 nm, so ist die Photosyntheseleistung wieder hoch. Diesen Steigerungseffekt nennt man nach seinem Entdecker Emerson-Effekt. Dieses scheinbar paradoxe Ergebnis wird erklärbar, wenn man die Existenz zweier Photosysteme mit einem jeweils spezifischen Reaktionszentrum annimmt. Das Reaktionszentrum des Photosysterns 1 ist das Chlorophyll a, oder Pigment 700. das Chlorophyllmolekül im Reaktionszentrum des Systems II wird Chlorophyll aII oder Pigment 682 genannt. Im Spektralbereich unterhalb von 682 nm absorbieren beide Photosysteme in einem ausgewogenen Verhältnis zueinander. Im Bereich oberhalb von 682 nm dominiert das System I über das System Il. Die Photosynthese wird dort durch den Mangel an Produkten des Photosystems, II begrenzt. Dies führt zu einer Leistungsverminderung des Gesamtsystems. Im EMERSON-Experiment wird diese Schwäche durch die zusätzliche Belichtung mit der kürzeren Wellenlänge aufgehoben. Bilder: Kroll, Demmer, Thies: "Stoffwechsel", Serie Biologie, Westermann Wien. |
|
|
Welt der Biologie |
| Anatomie/Physiologie | Botanik | Cytologie | Evolution | |
| Genetik | Humanbiologie | Ökologie | Sexualbiologie | Zoologie |
| Geschichte | Texte, Referate | Sehenswert | Kontakt | Physik |
Zwei Genetikthriller vom Autor dieser Seite |
.png)
Rudolf Oeller:Typhons RacheThriller über die vernichtende Kraft der Rache und den Traum vom ewigen Leben im Diesseits.
|
.png)
Rudolf Oeller:Typhon DistrictThriller über eine Gruppe junger Wissenschaftler, die an ihrer Gier zugrunde ging.
|
 Um die
physiologischen Abläufe der Photosynthese noch genauer zu verstehen,
muss man zu den physikalischen und chemischen Grundlagen des Photosyntheseprozesses
vordringen. Dieser spielt sich im wesentlichen in den molekularen Strukturen
der Chloroplasten ab.
Um die
physiologischen Abläufe der Photosynthese noch genauer zu verstehen,
muss man zu den physikalischen und chemischen Grundlagen des Photosyntheseprozesses
vordringen. Dieser spielt sich im wesentlichen in den molekularen Strukturen
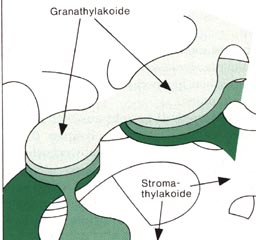
der Chloroplasten ab. Der Chloroplast
ist durch eine Doppelmembran gegen das Plasma abgegrenzt. Der Innenraum
wird von einem Membrankörper durchzogen, der in die feingranuläre
Grundsubstanz eingebettet ist Dieses Membransystem ist in zwei Bereiche
gegliedert Die abgeflachten Doppelmembranen (Thylakoide) erscheinen an
manchen Stellen wie ein Stapel flacher Scheiben.
Der Chloroplast
ist durch eine Doppelmembran gegen das Plasma abgegrenzt. Der Innenraum
wird von einem Membrankörper durchzogen, der in die feingranuläre
Grundsubstanz eingebettet ist Dieses Membransystem ist in zwei Bereiche
gegliedert Die abgeflachten Doppelmembranen (Thylakoide) erscheinen an
manchen Stellen wie ein Stapel flacher Scheiben.