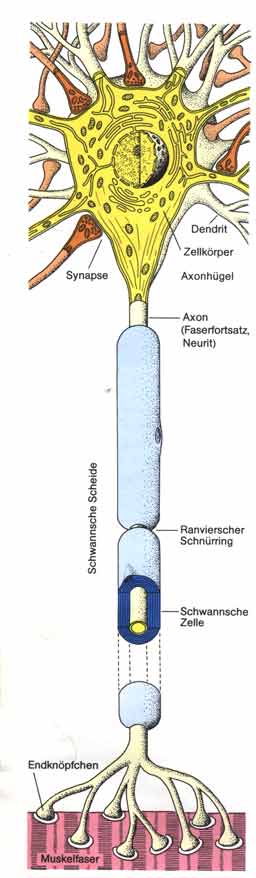
 Bau
einer Nervenzelle Bau
einer Nervenzelle
Ein 5 mm2 großes, 0,007 mm dickes, angefärbtes
Stück aus der Großhirnrinde wird untersucht. Im Lichtmikroskop
erkennt man in diesem mikroskopischen Präparat eine gleichförmige,
homogene Grundmasse, in die zahlreiche Körner eingebettet sind. Diese
Körner sind die von Cytoplasma umgebenen Zellkerne von Nervenzellen.
Den Zellkern mit dem umgebenden Cytoplasma nennt man Zellkörper.
Mit den üblichen Färbemethoden heben sich die Nervenzellen,
auch Neurone genannt, kaum voneinander ab. Mit Metallsalzen, beispielsweise
einer Silbernitratlösung, lassen sich dagegen einzelne Zellen schwarz
imprägnieren. Mit feinen Mikropipetten kann man Farbstoff in einzelne
Zellkörper injizieren. Die Farbe verteilt sich dann über die
ganze Zelle und hebt sie so von den umliegenden Neuronen ab. Die Unterschiede
zwischen verschiedenen Nervenzelltypen sind erheblich. Entscheidend sind
jedoch ihre Gemeinsamkeiten:
- Der Zellkörper enthält den Zellkern und den größten
Teil des Cytoplasmas.
- Vom Zellkörper der Nervenzellen gehen zahlreiche,
bäumchenartig verzweigte Fortsätze aus, die Dendriten.
- An einer Stelle verläßt ein langer Faserfortsatz,
das Axon, den Zellkörper. An seinem Ende verzweigt sich das Axon.
Jede Verzweigung endet in einer kleinen Verdickung, dem Endknöpfchen.
- Oft sind die Axone von vielen hintereinander liegenden
Zellen umwickelt. Diese Isolationshülle nennt man nach ihrem Entdecker
Schwannsche Scheide, die Lücken zwischen den Schwannschen Zellen
Ranviersche Schnürringe. Axone mit Schwannscher Scheide werden als
markhaltig bezeichnet, Axone ohne diese Hülle sind marklos.
Ruhepotential und Aktionspotential
Wird eine Elektrode außen am Axon angelegt und die andere ins Innere
eingestochen, zeigt das Oszilloskop eine Spannung von 60 - 90 mV an. Die
Innenseite ist negativ gegenüber der Außenseite. Vereinbarungsgemäß
wird immer das Vorzeichen der Innenseite angegeben.
• Am ungereizten Axon besteht zwischen außen und innen eine
Spannung von -60 bis -90 mV. Sie wird als Ruhepotential der Nervenzelle
bezeichnet.
Abbildung: Ruhepotentiale und Aktionspotentiale, wie sie sich von
Axonen ableiten und mit dem Oszilloskop sichtbar machen lassen.
Mit dem Oszilloskop kann man die elektrischen Impulse sichtbar machen,
die nach Reizung über das Axon laufen. Beim Durchgang eines Impulses
durch die Messstelle nimmt das Ruhepotential zunächst bis auf Null
ab. Dann wird das Innere des Axons positiv, und die Spannung steigt je
nach Neuron rasch auf + 20 bis + 40 mV an. Die Spannungsänderung
beträgt also insgesamt etwa 100 mV. Bereits 0,5 - 1 ms später
ist das Ruhepotential wiederhergestellt. Der elektrische Impuls hat die
Messstelle durchlaufen.
• Die Impulse von 100 mV, die über das Axon laufen, werden
als Aktionspotentiale bezeichnet.
• Die Leitung von Aktionspotentialen über die Axone nennt
man Erregungsleitung.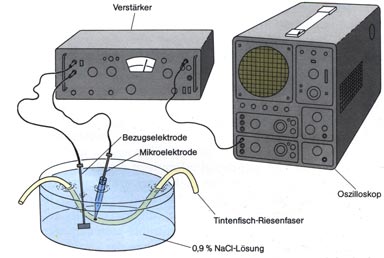
Strenggenommen sind Ruhepotential und Aktionspotentiale "Potentialdifferenzen"
oder Spannungen.
Abbildung:
Versuchseinrichtung zur Ableitung von Ruhepotentialen und Aktionspotentialen
bei Riesenfasern.
Ionenverteilung: In elektrisch leitenden Drähten wandern
Elektronen vom negativen zum positiven Pol. In wässeriger Umgebung,
wie in Nervenzellen, kann elektrische Ladung nicht in Form von Elektronen
fließen.
Untersucht man Nervenzellen, stellt man dort viele frei bewegliche
Ionen fest.
Axone sind außen und innen elektrisch neutral; es stehen sich
jeweils gleich viele positiv geladene Kationen und negativ geladene Anionen
gegenüber. Betrachtet man aber die Verteilung der einzelnen Ionenarten
außerhalb und innerhalb der Membran, so stellt man große Unterschiede
fest: Proteinanionen kommen außen nicht vor. Die Kaliumionenkonzentration
ist innen etwa 50mal höher als außen. Für die Chloridionen
gilt das Umgekehrte. Die Natriumionenkonzentration ist außen 10mal
höher als innen. Diese Verhältnisse gelten für das Ruhepotential.
Entstehung einer Gleichgewichtsspannung:
Lassen sich mit Ionen Spannungen erzeugen und durch lebende Zellen weiterleiten?
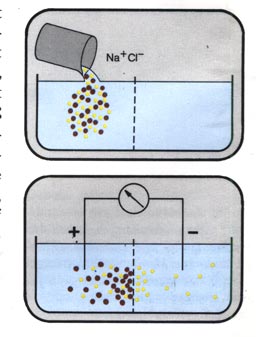
Dazu ein Versuch (siehe Abbildung rechts):
Gießt man in ein Gefäß mit Wasser auf einer Seite eine
Natriumchloridlösung (Na+Cl-), verteilen sich
die Na+-Ionen und Cl--Ionen im Laufe der Zeit durch
Diffusion gleichmäßig in der Flüssigkeit. Ist das Gefäß
durch eine selektiv permeable Membran in zwei Hälften getrennt, die
zwar die kleineren CI--Ionen, nicht aber die Na+
Ionen passieren läßt, gelangen nur CI--Ionen in
die andere Hälfte. Da sich entgegengesetzt geladene Ionen anziehen,
wandern Na+-Ionen bis zur Membran mit. Das wirkt sich auf die
CI--Ionen aus: CI--Ionen gelangen nur so lange zur
anderen Seite, bis der „Druck" durch das Konzentrationsgefälle
so groß ist wie der „Zug" durch die Na+-Ionen.
Ein Gleichgewicht stellt sich ein. Die Kraft, die den Ausstrom zum Erliegen
bringt, ist als Gleichgewichtsspannung messbar.
Entstehung des Ruhepotentials: Auch Axone haben eine selektiv
permeable Membran, die Axonmembran. Sie enthält viele Kanäle,
die für K+-Ionen durchlässig sind. Wegen des Konzentrationsgefälles
diffundieren K+-Ionen durch die Kanäle nach außen.
Die Protein-Ionen sind so groß, dass sie nicht durch die Membran
diffundieren können. Sie üben aber, da sie negativ geladen sind,
einen Zug auf die K+-Ionen aus. Je mehr K+-Ionen
nach außen diffundieren, desto stärker wird der Zug von innen.
Nach einiger Zeit stellt sich ein Gleichgewicht zwischen Konzentrationsgefälle
und elektrischer Anziehungskraft ein. Die dabei herrschende Spannung
ist das Ruhepotential. Das Ruhepotential entspricht praktisch der
Gleichgewichtsspannung für K+-Ionen.
• Das Ruhepotential ist vor allem ein Kaliumdiffusionspotential.
Natrium-Kalium-Pumpe:
In Wirklichkeit strömen nicht nur K+-Ionen, sondern auch
Na+- und CI--Ionen durch die Membran, allerdings
in viel geringerem Maße. Daher haben sie auch kaum Einfluss auf
das Ruhepotential. Versuche mit radioaktiven 42K+-Ionen
und 24Na+-Ionen ergaben, dass ständig eindringende
Na+-Ionen unter Energieverbrauch nach außen gepumpt werden.
Zugleich gelangt die gleiche Menge K+-Ionen nach innen. Man spricht deshalb
von der Natrium-Kalium-Pumpe.
Entstehung der Aktionspotentiale:
Über Elektroden, an die man eine variable Spannung anlegt, lässt
sich das Ruhepotential der Axone gezielt verändern. Fließt
von der inneren Elektrode ein schwacher Strom durch die Membran hindurch
zur äußeren Elektrode, nimmt das Ruhepotential ab. Dies
nennt man Depolarisation. Fließt kein Strom zwischen den Elektroden,
wird das Ruhepotential wieder aufgebaut. Wird aber stärker depolarisiert,
so dass das Ruhepotential auf einen Wert von etwa -50 mV absinkt, wird
schlagartig ein Aktionspotential ausgelöst.
Versuche mit 42K+ und 24Na+
ergaben: Nimmt das Ruhepotential durch Depolarisation ab, werden in der
Membran immer mehr Natriumkanäle geöffnet. Dadurch gelangen
zunehmend mehr Na+-Ionen nach innen. Dies läßt das
Ruhepotential weiter abnehmen, noch mehr Kanäle werden geöffnet.
Die Spannung geht schließlich auf 0 zurück, überschreitet
die Nullmarke und erreicht einen Wert von 20 bis 40 mV. Die Gleichgewichtsspannung
von +60 mV für Na+ wird also nicht ganz erreicht. Der
Grund dafür: Schon kurze Zeit vorher beginnt die Durchlässigkeit
der Membran für Na+ wieder abzunehmen, für K+
dagegen nimmt sie zu. Jetzt strömen K+-Ionen nach außen,
das Aktionspotential geht wieder zurück. Schließlich ist nach
0,5-1 ms die Ruhespannung wieder hergestellt.
• Aktionspotentiale entstehen, wenn die Axonmembran stark genug
depolarisiert wird. Dann nimmt ihre Durchlässigkeit für Natriumionen
für kurze Zeit auf über das Hundertfache des Ruhewertes zu.
Durch die Natrium-Kalium-Pumpe werden K+ und Na+
im Laufe der Zeit wieder zurücktransportiert. Man hat gemessen, dass
pro Aktionspotential und pro cm2 Axonoberfläche nur 3
- 10-12 Mol Na+ und etwas weniger K+
durch die Membran gelangen. Daraus ergibt sich, dass auch ohne Zurückpumpen
Tausende von Aktionspotentialen entstehen könnten.
Kontinuierliche und saltatorische Erregungsleitung
Marklose Axone:
Wo in einem Axon ein bereits erregter Bereich an einen noch unerregten
Bereich grenzt, stoßen sowohl innen als auch außen an der
Membran entgegengesetzt geladene Zonen aufeinander. In diesem Grenzbereich
beginnen die Ionen zu wandern; zunächst innen und außen der
Membran entlang, dann auch durch diese hindurch. Die Membran wird so stark
depolarisiert, dass ein neues Aktionspotential ausgelöst wird. Ein
neuer Grenzbereich bildet sich, wiederum wandern Ionen, die Membran wird
depolarisiert, neue Aktionspotentiale werden ausgelöst. Fortschreitende
Depolarisation führt also zum kontinuierlichen Fortschreiten von
Aktionspotentialen über das Axon.
Wird im Versuch ein Axon in der Mitte gereizt, so kommt es zu einer Ausbreitung
der Depolarisation und damit der gebildeten Aktionspotentiale in beide
Richtungen. Im Nervensystem tritt dieser Fall nicht auf. Dort laufen die
Aktionspotentiale stets in einer Richtung über das Axon. Eine Umkehr
der Richtung ist nicht möglich, weil an den Stellen, an denen eben
noch ein Aktionspotential ausgebildet war, die
Membran 1 ms lang überhaupt nicht und weitere 1-2 ms lang kaum errembar
ist. Diese Zeitspanne wird als Refraktärzeit bezeichnet.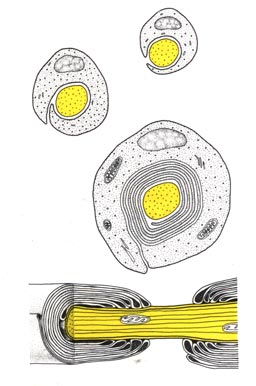
Wie rasch marklose Axone die Erregung leiten, hängt von ihrem Querschnitt
ab. Die 1µm dicken marklosen Schmerzfasern der Katze erreichen 1
m/s, die 500 µm dicken Riesenfasern der Tintenfische 20m/s.
Abbildung: Eine Zelle legt sich an das Axon und wickelt sich herum.
So entsteht eine isolierende Hülle, die "Schwannsche Scheide".
Der Spalt zwischen zwei Schwannschen Zellen ist der "Ranviersche
Schnürring".
Markhaltige Axone:
Laufen Aktionspotentiale über markhaltige Axone, spielen sich dieselben
Vorgänge ab. Weil die Schwannschen Zellen aber eine dichte Isolationshülle
bilden, können nur an den Schnürringen Aktionspotentiale entstehen.
Die Strecke dazwischen, je nach Axon sind das 1 - 5 mm, wird von den Ionen
„überbrückt", die zum Ladungsausgleich innen an der
Axonmembran und außen an den Schwannschen Zellen entlangwandern.
Die Aktionspotentiale laufen also nicht kontinuierlich über das
Axon, sondern springen von Schnürring zu Schnürring. Diese Art
der Erregungsleitung geht rascher: Markhaltige Axone leiten bei einem
Durchmesser von 10-20 µm die Erregung mit Geschwindigkeiten bis
130 m/s.
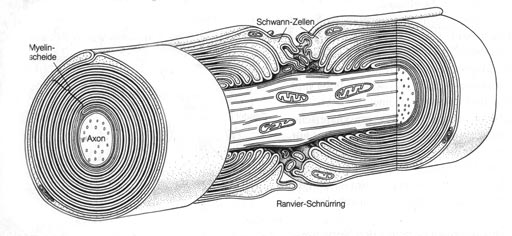
Abbildung: Nervenfaser mit Myelinscheide und Ranvier-Schnürring
(schematischer Längs- und Querschnitt)
(aus. G. Czihak et al. "Biologie", Springer)
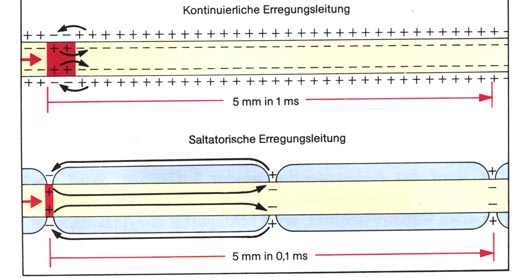
Abbildung:
• Bei marklosen Axonen laufen die Aktionspotentiale kontinuierlich
über das Axon:
kontinuierliche Erregungsleitung.
• Bei markhaltigen Axonen springen die Aktionspotentiale von Schnürring
zu Schnürring:
saltatorische Erregungsleitung.
Bilder:
Goll/Schwoerbel: Sinne, Nerven, Hormone (Cornelsen-Velhagen & Klasing)
und
G. Czihak etc. al "Biologie" (Springer)
|




.png)
.png)