
|
| |
|
|
CYTOLOGIE |
Meiose (ausführlich) |
|
|
Download: |

|
|
|
Meiose(Ausführliche Version) Die Meiosis ist im
Gegensatz zur Mitose keine erbgleiche Teilung. Die Meiose besteht aus zwei aufeinanderfolgenden Teilungen, der ersten und der zweiten meiotischen Teilung. Die für die Meiose typischen Vorgänge laufen in der ersten Teilung ab, während die zweite im Prinzip der Mitose entspricht. Wir wollen bei der Besprechung des meiotischen Stadienablaufs von der Situation bei diploiden Organismen ausgehen. In der Keimbahn höherer Tiere und des Menschen sowie in den Pollensäcken der höheren Pflanzen sorgen Mitosen zunächst für die Gesamtzahl derjenigen Zellen, die als Meiocyten später in die Meiose eintreten. In den Antheren liegt zwischen den letzten prämeiotischen Mitosen und dem Beginn der Meiose eine Periode von einigen Tagen, in denen keine mikroskopisch erkennbaren cytologischen Vorgänge ablaufen. Bei Lilien dauert die prämeiotische Replikation der DNA etwa 6mal länger als die S - Phase mitotischer Interphase - Kerne. Dann treten die Zellen, die sich bisher ausschließlich mitotisch geteilt haben, in die Meiose ein und durchlaufen damit einen prinzipiell andersartigen Teilungsvorgang. Über die biochemischen Vorgänge, die zu dieser Umstimmung der Zellen führen, ist noch nichts bekannt. Am Anfang der Meiose steht ein diploider Interphasekern. Er besitzt zwei einander entsprechende Chromosomensätze, zwei Genome, von denen eines ursprünglich aus der väterlichen, das andere aus der mütterlichen Keimzelle stammt. Jedem Chromosom des väterlichen Genoms entspricht ein bestimmtes Chromosom im mütterlichen Genom. Diese Chromosomen werden als Homologe bezeichnet; sie stimmen in ihrer Längsstruktur völlig überein. Die Homologie erstreckt sich auch auf ihren Gengehalt, sie ist im Interphasekern mikroskopisch nicht erkennbar. Die Chromosomen sind in diesem Stadium an der Kernmembran angeheftet. Prophase I
|
|
|
Leptotän, Zygotän, Pachytän, Diplotän und Diakinese.Leptotän
Zygotän Pachytän Im Pachytän kommt es zu Austauschvorgängen zwischen einander entsprechenden Abschnitten der Chromatiden. Sie führen zu einem der wichtigsten Effekte der Meiose, zum Umbau der Chromatiden der beiden homologen Chromosomen. Als Folge kommt eine Neukombination der Gene der elterlichen Genome zustande. Diese Vorgänge werden als Crossing over bezeichnet. Die Überkreuzungsstellen zwischen den Chromatiden, die Chiasmata, sind bei günstigen Objekten im Diplotän mikroskopisch sichtbar. Zwischen Beginn und Ende der Paarung - also vorn Zygotän bis zum frühen Diplotän - ist im Elektronenmikroskop zwischen den gepaarten Homologen eine dreiteilige Struktur sichtbar, die als synaptischer oder synaptonemaler Komplex bezeichnet wird. Beiderseits eines Zentralelements verlaufen parallel 2 Lateralelemente. Er ist für die Bivalentenbildung erforderlich und hält die beiden Homologen in einem Abstand von etwa 0,10 mm. Die Initiation kann an vielen Stellen der Homologen gleichzeitig beginnen; im Pachytän besitzt jedes Bivalent über seine gesamte Länge hinweg einen kontinuierlichen Komplex. Homologenpaarung und Anwesenheit des synaptischen Komplexes sind Voraussetzungen für den Ablauf von Crossing - over - Vorgängen und damit für den Genaustausch zwischen den Homologen. Die engen Beziehungen zwischen diesen beiden Phänomenen gehen aus der Tatsache hervor, dass der synaptische Komplex bei denjenigen Organismen fehlt, bei denen keine Rekombinationsvorgänge ablaufen. Das gilt z. B. für Drosophila - Männchen. Im Zygotän und Pachytän sind im Bereich des synaptischen Komplexes sogenannte "recombination nodules" erkennbar. Es handelt sich um knotenförmige Gebilde der Chromatiden, die sich an das Zentralelement des Komplexes anheften. Sie sind zufällig über die Länge der Bivalente verteilt; ihre Anzahl ist mit der Zahl und Verteilung der Crossing - over - Punkte bzw. der Chiasmata korreliert. Ihre Funktion besteht möglicherweise darin, die für das Crossing over notwendigen molekularen Prozesse zu vermitteln oder durchzuführen. Diplotän Diakinese Die auf das Diplotän folgenden Stadien sind im wesentlichen durch zwei Vorgänge charakterisiert: Der Kontraktionsgrad der Chromosomen nimmt stark zu, außerdem rücken die Chiasmata bei der Mehrzahl aller Arten an die Chromosomenenden. Die Mechanik dieses Prozesses ist noch nicht geklärt. Ist es in beiden Armen zur Chiasmenbildung gekommen, so entstehen geschlossene Bivalente; falls nur ein Arm Chiasmata besitzt, so sind die Bivalente offen. Chiasmenausfall führt zur Bildung von Univalenten. Bei einigen Objekten treten als konstantes Artmerkmal interstitielle Chiasmata auf. Sie terminalisieren nicht, sondern verbleiben am Ort ihrer Entstehung. Beim Übergang von der späten Diakinese zur ersten Metaphase laufen im Prinzip die gleichen Vorgänge ab wie im entsprechenden Stadium der Mitose: Kernmembran und Nucleolen werden aufgelöst, und es wird eine Spindel gebildet, deren Elemente mit den Centromeren der Chromosomen in Verbindung treten. Damit ist die 1. meiotische Prophase abgeschlossen. Metaphase I Anaphase I Telophase I
Interkinese Die Chromosomen lockern sich auf. Kernhüllen und Nucleolen werden gebildet. Es entstehen also Interphase - Kerne, es erfolgt jedoch keine Replikation der DNA. Jeder der beiden haploiden Tochterkerne enthält zwar ein vollständiges Genom, die Genome sind bei heterozygoten Organismen in ihrer genetischen Zusammensetzung jedoch nicht identisch. Auf die erste meiotische Teilung folgt ohne ein längeres Zwischenstadium die zweite Teilung. Sie verläuft nach den Gesetzmäßigkeiten der Mitose und reproduziert diejenige cytologische und genetische Situation, die am Ende der ersten Teilung realisiert war. |
|
|
Prophase II Die Chromosomen kondensieren sich erneut. In der späten Prophase werden Kernmembran und Nucleolen aufgelöst, und es werden Kernspindeln gebildet.
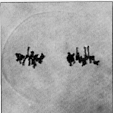 Metaphase II
Metaphase II
Die Chromosomen werden in die Metaphaseplatten eingeordnet. In der Mikrosporogenese und der Spermatogenese führt jeder der beiden haploiden Kerne die Teilung unabhängig vom anderen Kern durch. In der Meiocyte sind folglich zwei Äquatorialplatten mit zwei Kernspindeln vorhanden.
Aus den Metaphaseplatten wandern Chromatiden polwärts.
Die Chromatiden sind an den Polen angekommen. In der diploiden Mutterzelle sind nunmehr 4 haploide Chromosomengruppen vorhanden. Sie werden teils durch Elemente der Spindel, teils durch ein neues Zellorganell, den Phragmoplasten, in einer optimalen gegenseitigen Lage festgehalten. Interphase II Die 4 Chromosomengruppen wandeln sich zu 4 Interphasekernen um. Die Chromatiden dekondensieren sich, Kernmembranen und Nucleolen werden neu gebildet. Damit ist der Stadienablauf der Meiose beendet. Durch Einzug von Zellwänden wird das Cytoplasma der Mutterzelle den vier Kernen zu etwa gleichen Teilen zugeordnet. Auf diese Weise entstehen in der H&u uml;lle der Mutterzelle 4 selbständige haploide Zellen, die Gonen. Bei den Diplonten sind sie die Vorläufer der Keimzellen und entwickeln sich bei verschiedenen Organismengruppen in etwas unterschiedlicher Weise zu den Gameten weiter. |
 |
Vorarlberger Bildungsserver |
| Anatomie/Physiologie | Botanik | Cytologie | Evolution | |
| Genetik | Humanbiologie | Ökologie | Sexualbiologie | Zoologie |
| Geschichte | Texte, Referate | Sehenswert | Kontakt | Physik |
Zwei Genetikthriller vom Autor dieser Seite |
.png)
Rudolf Oeller:Typhons RacheThriller über die vernichtende Kraft der Rache und den Traum vom ewigen Leben im Diesseits.
|
.png)
Rudolf Oeller:Typhon DistrictThriller über eine Gruppe junger Wissenschaftler, die an ihrer Gier zugrunde ging.
|
 Es ist nicht angebracht, den Begriff "Reduktionsteilung"
für den Gesamtvorgang zu verwenden. Die Reduktion der Chromosomenzahl
ist nur einer der Effekte der Meiose, in genetischer Beziehung der am
wenigsten bedeutsame. Viel wichtiger sind die Konsequenzen, die sich aus
der Umgestaltung der Chromosomen und Genome im Zuge der Meiose ergeben.
Es ist nicht angebracht, den Begriff "Reduktionsteilung"
für den Gesamtvorgang zu verwenden. Die Reduktion der Chromosomenzahl
ist nur einer der Effekte der Meiose, in genetischer Beziehung der am
wenigsten bedeutsame. Viel wichtiger sind die Konsequenzen, die sich aus
der Umgestaltung der Chromosomen und Genome im Zuge der Meiose ergeben. In der ersten meiotischen Prophase kommt eine Kondensation der Chromosomen
zustande, die in der Regel zu einem beträchtlich stärkeren Kontraktionsgrad
als in der Mitose führt. Die wesentlichsten Vorgänge der Meiose
laufen in diesem Abschnitt ab. Man hat ihn deshalb in eine Serie aufeinanderfolgender
Stadien unterteilt, für die es in der Mitose keine Parallelen gibt.
Hierbei unterscheidet man:
In der ersten meiotischen Prophase kommt eine Kondensation der Chromosomen
zustande, die in der Regel zu einem beträchtlich stärkeren Kontraktionsgrad
als in der Mitose führt. Die wesentlichsten Vorgänge der Meiose
laufen in diesem Abschnitt ab. Man hat ihn deshalb in eine Serie aufeinanderfolgender
Stadien unterteilt, für die es in der Mitose keine Parallelen gibt.
Hierbei unterscheidet man: Aus dem Chromatinknäuel des Interphasekerns differenzieren sich optisch
die Chromosomen als lange, dünne Fäden heraus, Obwohl sie bereits
in ihre Chromatiden unterteilt sind, wird ihre Doppelnatur im Lichtmikroskop
nicht erkennbar.
Aus dem Chromatinknäuel des Interphasekerns differenzieren sich optisch
die Chromosomen als lange, dünne Fäden heraus, Obwohl sie bereits
in ihre Chromatiden unterteilt sind, wird ihre Doppelnatur im Lichtmikroskop
nicht erkennbar.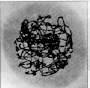 Der wichtigste Vorgang während der frühen Stadien der ersten
meiotischen Prophase besteht in der äußerst exakten Paarung
der homologen Chromosomen. Sie beginnt oftmals an den Chromosomenenden,
die bei vielen Objekten an der Kernmembran angeheftet sind, und schreitet
in Richtung auf das Centromer fort. Über die Mechanik der Paarung
liegen noch keine gesicherten Befunde vor. Sie kommt mit größter
Präzision nur zwischen homologen Chromosomenregionen - d. h. zwischen
identischen oder einander entsprechenden Genen - zustande. Wird die Längsstruktur
der Chromosomen durch Chromosomenmutationen verändert, so läuft
die Paarung nur zwischen homologen Abschnitten ab. Als Folge dieser Gesetzmäßigkeit
werden in den frühen meiotischen Stadien von Organismen, die heterozygot
für Chromosomenmutationen sind, komplizierte Paarungsfiguren gebildet.
Inhomologe Paarung kommt gelegentlich in der Meiosis haploider Organismen
oder bei Artbastarden zustande.
Der wichtigste Vorgang während der frühen Stadien der ersten
meiotischen Prophase besteht in der äußerst exakten Paarung
der homologen Chromosomen. Sie beginnt oftmals an den Chromosomenenden,
die bei vielen Objekten an der Kernmembran angeheftet sind, und schreitet
in Richtung auf das Centromer fort. Über die Mechanik der Paarung
liegen noch keine gesicherten Befunde vor. Sie kommt mit größter
Präzision nur zwischen homologen Chromosomenregionen - d. h. zwischen
identischen oder einander entsprechenden Genen - zustande. Wird die Längsstruktur
der Chromosomen durch Chromosomenmutationen verändert, so läuft
die Paarung nur zwischen homologen Abschnitten ab. Als Folge dieser Gesetzmäßigkeit
werden in den frühen meiotischen Stadien von Organismen, die heterozygot
für Chromosomenmutationen sind, komplizierte Paarungsfiguren gebildet.
Inhomologe Paarung kommt gelegentlich in der Meiosis haploider Organismen
oder bei Artbastarden zustande.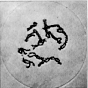 Wenn die Parallelkonjugation der homologen Chromosomen vollzogen ist,
befindet sich der Kern im Pachytän, einem der wichtigsten Stadien
der Meiosis. Die Chromosomen liegen nicht mehr in Form von Einzelelementen,
sondern von homologen Zweiergruppen, von Bivalenten, vor. Da jedes Chromosom
aus 2 Chromatiden besteht, stellt das Bivalent eine Vierergruppe, eine
Tetrade von strukturell selbständigen Längseinheiten dar.
Die Kondensation ist so weit fortgeschritten, dass die Pachytänbivalente
bei günstigen Objekten analysierbar und identifizierbar sind. Mit
Hilfe von Pachytänanalysen lassen sich evolutionistische Fragestellungen
bearbeiten, außerdem lassen sich die Folgen von Chromosomenmutationen
analysieren.
Wenn die Parallelkonjugation der homologen Chromosomen vollzogen ist,
befindet sich der Kern im Pachytän, einem der wichtigsten Stadien
der Meiosis. Die Chromosomen liegen nicht mehr in Form von Einzelelementen,
sondern von homologen Zweiergruppen, von Bivalenten, vor. Da jedes Chromosom
aus 2 Chromatiden besteht, stellt das Bivalent eine Vierergruppe, eine
Tetrade von strukturell selbständigen Längseinheiten dar.
Die Kondensation ist so weit fortgeschritten, dass die Pachytänbivalente
bei günstigen Objekten analysierbar und identifizierbar sind. Mit
Hilfe von Pachytänanalysen lassen sich evolutionistische Fragestellungen
bearbeiten, außerdem lassen sich die Folgen von Chromosomenmutationen
analysieren. Die Homologen zeigen die Tendenz, sich wieder voneinander zu trennen,
sie werden jedoch durch die Chiasmata zusammengehalten, die als Chromatinbrücken
zwischen den Nichtschwester - Chromatiden aufzufassen sind. Bei günstigen
Objekten sind sie sichtbar. Ihre Anzahl variiert in Abhängigkeit
von der Chromosomenlänge zwischen 1 und 8; die meisten Bivalente
haben 2 bis 3 Chiasmata. Die Kondensation der Chromosomen nimmt zu; der
Kern wird übersichtlicher. Trotzdem ist das Diplotän nur bei
wenigen Arten eindeutig analysierbar.
Die Homologen zeigen die Tendenz, sich wieder voneinander zu trennen,
sie werden jedoch durch die Chiasmata zusammengehalten, die als Chromatinbrücken
zwischen den Nichtschwester - Chromatiden aufzufassen sind. Bei günstigen
Objekten sind sie sichtbar. Ihre Anzahl variiert in Abhängigkeit
von der Chromosomenlänge zwischen 1 und 8; die meisten Bivalente
haben 2 bis 3 Chiasmata. Die Kondensation der Chromosomen nimmt zu; der
Kern wird übersichtlicher. Trotzdem ist das Diplotän nur bei
wenigen Arten eindeutig analysierbar.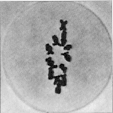 Im Gegensatz zur Mitose werden in der ersten meiotischen Metaphase nicht
Chromosomen, sondern Bivalente in die Metaphasenplatte eingeordnet. Vom
späten Zygotän bis zur 1 . Metaphase ist das Bivalent die Funktionseinheit.
Dies ist bei der mikroskopischen Bearbeiturig insofern vorteilhaft, als
man mit der haploiden Anzahl chromosomaler Einheiten arbeiten kann. Die
Chromosomen haben bei vielen Objekten eine nahezu würfelförmige
Gestalt und lassen keine Strukturmerkmale mehr erkennen. Es ist daher
in diesem Stadium nicht möglich, Karotyp - Analysen anzufertigen.
Die Orientierung der Homologen eines jeden Bivalentes innerhalb der Spindel
erfolgt zufallsgemäß. Dies führt zu einer weiteren Vermischung
des väterlichen und mütterlichen Erbguts in den entstehenden
Tochterkernen. Man bezeichnet diesen Effekt der Meiose als Umordnung der
Genome; er hat wichtige genetische Konsequenzen.
Im Gegensatz zur Mitose werden in der ersten meiotischen Metaphase nicht
Chromosomen, sondern Bivalente in die Metaphasenplatte eingeordnet. Vom
späten Zygotän bis zur 1 . Metaphase ist das Bivalent die Funktionseinheit.
Dies ist bei der mikroskopischen Bearbeiturig insofern vorteilhaft, als
man mit der haploiden Anzahl chromosomaler Einheiten arbeiten kann. Die
Chromosomen haben bei vielen Objekten eine nahezu würfelförmige
Gestalt und lassen keine Strukturmerkmale mehr erkennen. Es ist daher
in diesem Stadium nicht möglich, Karotyp - Analysen anzufertigen.
Die Orientierung der Homologen eines jeden Bivalentes innerhalb der Spindel
erfolgt zufallsgemäß. Dies führt zu einer weiteren Vermischung
des väterlichen und mütterlichen Erbguts in den entstehenden
Tochterkernen. Man bezeichnet diesen Effekt der Meiose als Umordnung der
Genome; er hat wichtige genetische Konsequenzen.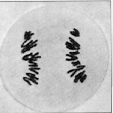 Jedes Bivalent trennt sich in seine beiden Homologen, die an entgegengesetzte
Spindelpole wandern. Wanderungseinheit ist also nicht die Chromaride,
sondern das Chromosom, wobei die beiden Chromatiden durch das gemeinsame
Centromer zusammengehalten werden. Hierin liegt einer der gravierendsten
Unterschiede zwischen Mitose und Meiosis. Als Konsequenz kommt die Reduktion
der Chromosomenzahl auf die Hälfte zustande.
Jedes Bivalent trennt sich in seine beiden Homologen, die an entgegengesetzte
Spindelpole wandern. Wanderungseinheit ist also nicht die Chromaride,
sondern das Chromosom, wobei die beiden Chromatiden durch das gemeinsame
Centromer zusammengehalten werden. Hierin liegt einer der gravierendsten
Unterschiede zwischen Mitose und Meiosis. Als Konsequenz kommt die Reduktion
der Chromosomenzahl auf die Hälfte zustande.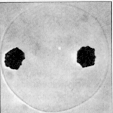 Die Wanderung der Chromosomen an die Spindelpole ist abgeschlossen.
In der ursprünglich diploiden Zelle sind zwei haploide Chromosomengruppen
vorhanden.
Die Wanderung der Chromosomen an die Spindelpole ist abgeschlossen.
In der ursprünglich diploiden Zelle sind zwei haploide Chromosomengruppen
vorhanden.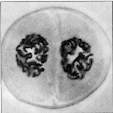
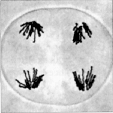 Anaphase II
Anaphase II 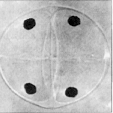 Telophase
II
Telophase
II