
|
|
||
|
|
EVOLUTIONSBIOLOGIE/SEXUALBIOLOGIE |
|
Vergleichende Embryogenese |
||
|
|
Siehe auch Bei Lurchen kommt es außerdem zur Ausbildung eines besonderen Larvenstadiums, der Kaulquappe, das eigentlich einem Embryonalstadium entspricht. Da es keinen Dot-tervorrat besitzt, muss es sich möglichst bald selbst ernähren. Kriechtiere und Vögel, die zum Landleben übergegangen sind
und deren Embryonal-entwicklung in einem trockenen Klima abläuft,
haben dieses Problem auf zwei Arten gelöst. Einmal sind ihre Eier
durch einen extrem hohen Dottervorrat ausgezeichnet. Die innere, durchsichtige Embryonalhülle wird als Amnion bezeichnet. Alle Wirbeltiere, die ein Amnion ausbilden, werden als Amnioten zusammengefasst. Es sind dies die Kriechtiere, die Vögel und die Säugetiere.
Die Embryonalentwicklung bei WirbellosenAbb.: Embryonalentwicklung der Insekten Im Gegensatz zum Rückenmark der Wirbeltiere ist für sie
ein Bauchmark typisch. Ein Neurulationsstadium im Sinne der Chordatiere
tritt bei Wirbellosen daher überhaupt nicht auf. Obwohl analoge Bildungen
bei den Wirbellosen und den Wirbeltieren antrifft, gibt es im wesentlichen
keine entwicklungsgeschichtlichen Entsprechungen zwischen den beiden Gruppen.
Am besten ist dies durch einen Vergleich der Insekten, die innerhalb der
Wirbellosen eine sehr hohe Evolutionsstufe einnehmen, und der Säuger
darzustellen. Die Embryonalentwicklung der Vögel und ReptilienNur die Dotterkugel mit der am animalen Pol sitzenden plasmatischen Keimscheibe entspricht der Eizelle des Vogels. Da die Dottermasse in den Furchungsprozess nicht einbezogen wird, bleibt die Embryogenese auf den kleinen Bereich der Keimscheibe beschränkt. Die etwa fünf Stunden nach der Befruchtung einsetzende Furchung liefert in 24 Stunden - solange verbleibt das Ei beim Huhn im Eileiter - ein der Blastula der Amphibien entsprechendes Keimstadium. Damit ist die Furchung abgeschlossen, und das Ei wird abgelegt. Für die Fortsetzung der Entwicklung ist eine Bebrütung notwendig.
Neurulation und Coelombildung vollzieht sich in der Folge wie bei den Amphibien. Wenn nach etwa 48 stündiger Bebrütung die Körpergrundgestalt des Embryos langsam sichtbar wird, beginnt die Bildung der Embryonalhüllen. Zu beiden Seiten des Keims wölben sich Ektoderm- und Mesodermepithel gemeinsam zu zwei Falten auf, die sich allmählich von vorn nach hinten über dem Embryo schließen. In dem so entstandenen Hohlraum, der Amnionhöhle (Fruchtblase), die sich mit einem eiweißreichen Sekret füllt, schwimmt der Embryo, geschützt vor Stößen und Austrocknung und von vier Zellschichten eingehüllt. Das transparente Innenblatt wird als Amnion bezeichnet, die derbe Außenschicht als Serosa. Die Entwicklung dieser Embryonalhüllen war sicher von größter Bedeutung beim Übergang der Wirbeltiere zum Landleben. Sie bleiben auch bei den Säugetieren erhalten, obwohl bei diesen die Embryonalentwicklung im Schutz des mütterlichen Körpers abläuft. Sie dienen bei diesen daher weniger dem Schutz des Embryos, sondern nehmen vielmehr engen Kontakt mit den Geweben des mütterlichen Organismus auf und werden zu Vermittlern des Stoffaustausches zwischen Embryo und Mutter. Im Verlauf der weiteren Entwicklung findet eine zunehmende Umwachsung des Dotters statt. Alle drei Keimblätter bilden dabei den Dottersack. Das Darmrohr bildet zuerst noch als offene Rinne den oberen Abschluss. Es dient allerdings in keinem Entwicklungsstadium der Aufnahme des Dotters. Diese Aufgabe erfüllen allein die dem Dotter aufliegenden Blutkapillaren des Dottersackes. Ein weiteres Problem stellt die Anhäufung von Stoffwechselschlacken im geschlossenen System des Eies dar. Nach etwa 60 stündiger Brutzeit bildet sich als Ausstülpung des embryonalen Enddarms ein embryonaler Harnsack, die Allantois. Sie übernimmt später auch die Funktion eines Atmungs- und Resorptionsorgans. In diesem Zusammenhang wird ein enger Kontakt mit der Eischale aufgenommen. Auf diesem Weg werden Calciumsalze aus der Schale resorbiert und dem Vogelembryo zum Aufbau seines Skelettes zugeführt. Ein Nebeneffekt ist die dabei eintretende Verdünnung der Schale, wodurch der spätere Schlüpfvorgang erleichtert wird. Interessant ist auch die Verwendung des Eiklars, das dem Embryo primär als Flüssigkeitsreserve dient – der Embryo trinkt sozusagen das Eiklar bis zum Schlüpfen. Die Embryonalentwicklung der Säugetiere und des Menschen
Die Eizellen der Säugetiere und die des Menschen sind durch einen Mangel an Reservestoffen gekennzeichnet. Wesentliche Unterschiede zur Entwicklung der Kriechtiere und Vögel ergeben sich sicher aus dem Umstand der Entwicklung im mütterlichen Organismus. So wurde vor allem die zeitliche Abfolge jener Entwicklungsschritte beschleunigt, die zur Bildung der Anlagen führen, die eine Verbindung zum mütterlichen Gewebe herstellen. Dies wird zum Beispiel durch den Verzicht auf zeitaufwendige Faltungsprozesse bei der Bildung der Amnionhöhle erreicht, die mit wenigen Ausnahmen (z. B. Hund, Katze) durch Hohlraumbildung inmitten des embryonalen Gewebes entsteht. Eine weitere Besonderheit der Säugerentwicklung ist die Ausbildung eines dem Stoffaustausch, der Ernährung und der Sauerstoffversorgung des Keims dienenden Organs, des Mutterkuchens oder der Plazenta. An seinem Aufbau sind sowohl embryonales als auch mütterliches Gewebe beteiligt. Auch die vergleichende Betrachtung dieser Anlage zeigt die entwicklungsgeschichtliche Abwandlung der Säuger von ihren Stammformen bis zum Menschen. Entsprechend der Ausbildung und Differenzierung einer Plazenta können Säugetiere in drei Unterklassen aufgeteilt werden. 1. Kloakentiere: In ihrer Entwicklung sehr ursprüngliche Säuger mit vielen reptilähnlichen Merkmalen, z. B. Kloake, Ablage weichhäutiger dotterreicher Eier. Hierher gehören wenige in Australien und Neuguinea lebende Arten wie der Ameisenigel und das Schnabeltier. 2 Beuteltiere: Vorwiegend auf Australien beschränkte Säuger mit doppelter Scheide und Gebärmutter und einer noch wenig ausgebildeten Plazenta. Sie gebären unentwickelte Junge, die aktiv über eine Schleimbahn in eine (an der Bauchseite der Weibchen liegende) Brusttasche (Beutel) kriechen, wo sie - an einer Zitze festgesaugt - ihre Entwicklung abschließen. Sie treten als Fleischfresser (zB. Beutelwolf, Beuteldachs, Beutelmaulwurf) oder Pflanzenfresser (Kängurus, Wombat) auf. 3. Plazentatiere: Zu ihnen gehören alle übrigen Säugetiere. Ihre Embryonalernährung erfolgt über eine gut entwickelte Plazenta in der Gebärmutter, in der die Jungen bis zum Abschluss ihrer Embryonalentwicklung verbleiben. Ein weiteres Kennzeichen ist der Besitz nur einer Scheide. Embryonalentwicklung des Menschen:Die an die Befruchtung anschließende Furchung läuft anfangs sehr langsam ab. So wird erst nach etwa 36 Stunden ein Zweizellenstadium und nach 80 Stunden ein Zwölfzel-lenstadium erreicht. Nach etwa sechs Tagen erreicht der Keim als so genannte Blastocyste die Gebärmutter, wo er sich meist im oberen Teil der Hinterwand einnistet. Dies erfolgt unter Mitwirkung von Enzymen, die die Gebärmutterschleimhaut und das darunterliegende Bindegewebe auflösen. In die so entstandene Höhlung sinkt der Keim ein und wird vom sich wieder schließenden Gebärmuttergewebe eingehüllt (Implantation). Die zur Blastocyste führenden Furchungsschritte sind durch zwei
Faktoren geprägt: - Erstens einem Furchungsmodus, der durch die Dotterarmut
des Säugereies bedingt ist und wie bei den dotterarmen Lurcheiern
abläuft. Bei Säugetieren übernehmen die den Hohlraum (Blastocystenhöhle)
begrenzenden Oberflächenzellen die Funktion der Ernährung des
Keims und werden dementsprechend als Trophoblast bezeichnet. An der Stelle,
an der die Blastocyste mit dem mütterlichen Gewebe in Berührung
kommt, ist ihre Wand verdickt. Aus dem Zellmaterial dieses Embryonalknotens
entwickelt sich der Embryo. Die dorsal (rückwärts) liegenden größeren Zellen, das primäre Ektoderm, und die ventral (bauchwärts) liegenden kleineren Zellen, das primäre Entoderm. Zwischen primärem Ektoderm und Trophoblast bildet sich die Amnionhöhle.
Um den Stoffaustausch und die Ernährung des Keims zu sichern,
kommt es zu einer besonderen Differenzierung des Trophoblasten. Seine
Oberfläche wird durch die Ausbildung eines reich verzweigten Zottennetzes
stark vergrößert. Diese Zotten werden bald von embryonalem
Bindegewebe und Blutgefäßen erfüllt. Der Trophoblast ist
damit zum Chorion (Zottenhaut) geworden. Es umgibt vorerst den ganzen
Keim, wird jedoch im Lauf der Entwicklung bis auf den Bereich des Haftstiels
abgebaut. Hier bildet es gemeinsam mit Teilen der Gebärmutterschleimhaut
den Mutterkuchen, die Plazenta. 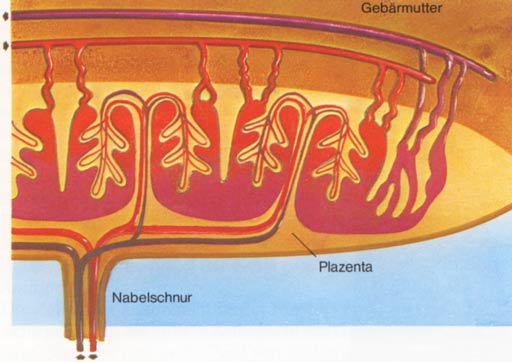 Von der Basalplatte ragen zahlreiche Septen in den Mutterkuchen und unterteilen ihn unvollkommen in etwa 15-30 Spalträume. In diese hängen von der Chorionplatte reich verästelte Zottenstämmchen hinein, die sowohl untereinander als auch mit den Septen und der Basalplatte verwachsen sind. Eine Ablösung ist hier also nicht möglich. Die Räume zwischen den Zotten sind von mütterlichem Blut erfüllt, das aus zahlreichen Arterien zuströmt, die Zotten umspült und am Rand der Plazenta durch die Gebärmuttervenen wieder abströmt. Das Blut des Keims wird in einem feinen Kapillarnetz durch das Zottennetz geführt. Die Blutkreisläufe von Mutter und Kind bleiben somit stets getrennt. Der Stofftransport innerhalb der Plazenta erfolgt nicht nur durch Diffusion oder Osmose, sondern es ist dabei auch eine aktive Teilnahme der Gewebe an den Grenzschichten zu beobachten. Ferner ist die Plazentaschranke nicht für alle Stoffe gleich gut durchlässig. Die Atmungsgase Sauerstoff und Kohlendioxid können entsprechend den Druckverhältnissen relativ leicht passieren. Nahrungsstoffe können im allgemeinen nur in ihren Grundbestandteilen (Einfachzucker; Aminosäuren, Glyzerin und Fettsäuren) übertreten. Verschiedentlich ist das Chorionepithel auch für Eiweißkörper, wie Antikörper oder Hormone, durchlässig. In gleicher Weise gelangen auch andere Antikörper von der Mutter in das Blut des Fetus und verleihen diesem einen ersten Immunitätsschutz. Auch vom Keim können Hormone in den Körper der Mutter gelangen. So kann zum Beispiel durch das den Blutzuckerspiegel regelnde Hormon Insulin des Fetus eine Zuckerkrankheit während der Schwangerschaft vorübergehend behoben werden. Darüber hinaus werden in der Plazenta zahlreiche, die Entwicklung und den Schwangerschaftsverlauf beeinflussende Hormone gebildet, die sowohl in den kindlichen wie in den mütterlichen Kreislauf gelangen. Gewissen Salzen gegenüber scheint die Plazenta hingegen nur in einer Richtung durchlässig zu sein. Calcium kann nur in Richtung Fetus passieren, der seinen Bedarf aus dem Körper der Mutter deckt. Auch Vitamine gelangen über die Plazenta zum Kind. Die Plazenta erfüllt Funktionen, die im erwachsenen Organismus Atmungsorgane,
Darm, Leber und Niere verrichten. Zusätzlich ist sie ein lebenswichtiges
biologisches Schutzorgan, das den Übertritt vieler schädlicher
Stoffe in den fetalen Kreislauf verhindert. Nach der Entstehung des Keimschildes läuft die Entwicklung in diesem
Bereich ähnlich wie bei den Kriechtiere und Vögel weiter. |
|
|
Vorarlberger Bildungsserver |
| Anatomie/Physiologie | Botanik | Cytologie | Evolution | |
| Genetik | Humanbiologie | Ökologie | Sexualbiologie | Zoologie |
| Geschichte | Texte, Referate | Sehenswert | Kontakt | Physik |
Zwei Genetikthriller vom Autor dieser Seite |
.png)
Rudolf Oeller:Typhons RacheThriller über die vernichtende Kraft der Rache und den Traum vom ewigen Leben im Diesseits.
|
.png)
Rudolf Oeller:Typhon DistrictThriller über eine Gruppe junger Wissenschaftler, die an ihrer Gier zugrunde ging.
|
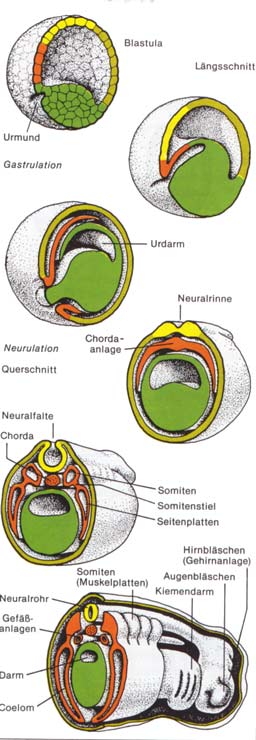 Abb.:
Embryonalentwicklung der Lurche
Abb.:
Embryonalentwicklung der Lurche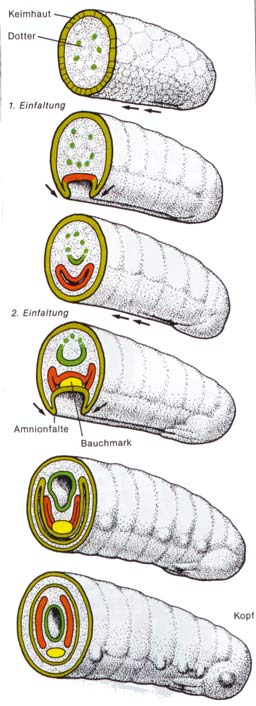 Die
Keimesentwicklung der wirbellosen Tiere ist dadurch gekennzeichnet, dass
die Anlage des Nervensystems sowie die ganze übrige Keimesdifferenzierung
von der Bauchseite ausgeht, bzw. vom vorderen Keimende (Kopf) nach hinten
hin fortschreitet.
Die
Keimesentwicklung der wirbellosen Tiere ist dadurch gekennzeichnet, dass
die Anlage des Nervensystems sowie die ganze übrige Keimesdifferenzierung
von der Bauchseite ausgeht, bzw. vom vorderen Keimende (Kopf) nach hinten
hin fortschreitet. 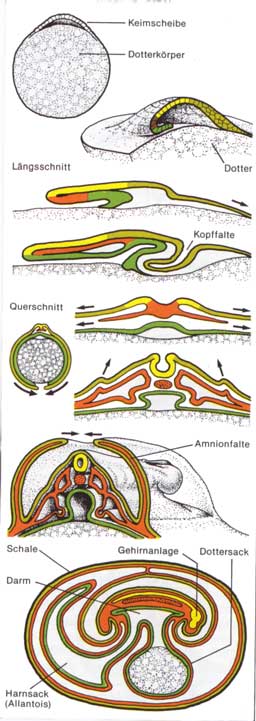 Abb.:
Embryonalentwicklung der Vögel und Reptilien
Abb.:
Embryonalentwicklung der Vögel und Reptilien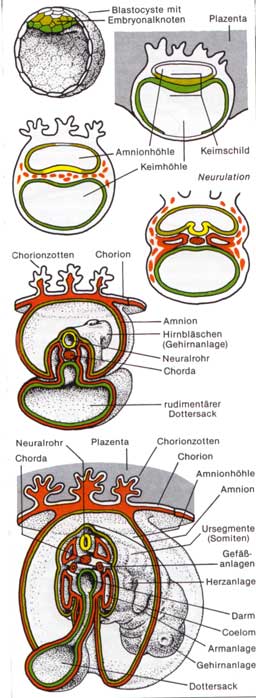 (unter
besonderer Berücksichtigung der Plazenta)
(unter
besonderer Berücksichtigung der Plazenta)